
Крымуков
Фамилия: Крымуков
Субэтнос: Кабардинец
Населенный пункт: Дыгулыбгей
Гаплогруппа: N1c >> Y3037
Y-хромосома, являющаяся основным компонентом данного исследования, уникальна тем, что передается строго по отцовской линии. Это позволяет проследить историю мужской линии.
Более 200 тыс. лет назад жил общий предок всех мужчин, так называемый Y-хромосомный Адам. Его потомство за это время распространилось по всему миру, разделяясь генетически на определённые ветви, которые учёными названы гаплогруппами. Время их возникновения десятки тысяч лет. За такой длительный период внутри гаплогрупп образовалось множество новых линий. Глубокий ДНК тест дает возможность выявить современную линию носителя.
Суть исследования не в определении национальности, однако его результаты могут продемонстрировать, насколько они характерны для конкретных регионов и возможно этносов. Строгая привязка определённых линий и гаплогрупп к каким-то народам очень сложная задача.
По результатам теста Крымукова N1c (или по современной номенклатуре N1a1). Линия предполагаемая у Крымукова чаще всего встречается в Восточной Азии (у китайцев и японцев). Также выявлена у татар. Среди других участников проекта совпадений нет, эта ветка крайне редко встречается на Кавказе.
Уникальная Y-хромосома
Y-хромосома уникальна тем, что передается строго по отцовской линии, что позволяет проследить историю мужской линии на протяжении многих поколений.
Глубокие (полногеномные) тесты позволяют получить наиболее подробную информацию о генетической линии. Если вы хотите получить максимум информации, рекомендуется сразу делать глубокий тест.
Исследование 37 и более маркеров позволяет провести сравнение с результатами тестов других участников и предсказать более глубокую линию происхождения. Эти тесты заметно дешевле, но и информацию дают намного меньше.
Важно отметить, что цель данного исследования не в определении национальности. Однако его результаты могут показать, насколько выявленные генетические линии характерны для конкретных регионов и, возможно, этносов.
В ходе анализа была определена гаплогруппа и более глубокий субклад. Эта информация помогает понять, к какой древней линии человечества принадлежит исследуемый образец.
Далее в публикации вы найдете подробную информацию о выявленной гаплогруппе, ее распространении, а также о возможных генетических связях с другими протестированными участниками проекта.
Для тех, кто хочет глубже разобраться в основах ДНК-генеалогии, на нашем сайте есть дополнительные образовательные материалы. Основные ссылки на эти материалы вы можете найти чуть ниже в этой публикации.
Также обратите внимание на ссылки на наши социальные сети. Нажав на них, вы можете подписаться и быть в курсе последних новостей проекта. Если у вас возникнут вопросы, наиболее оперативно вам ответят в нашем телеграм-чате.
- Рекомендуем посмотреть выступление Александра Ракитько. Примерно часовое выступление на тему Y-ДНК тестов. Ссылка на видео.
- Был ли у человечества один общий предок? // Дробышевский. Человек разумный
- ДНК-генеалогия как путешествие к истокам
- Термины и определения
- Гаплогруппа
- Как читать Y-древо
- Методика маркерного анализа
- Подробнее о древе Yfull
- Чем отличаются тесты Y хромосомы?
- Часто задаваемые вопросы
- Список протестированных фамилий
- Как заказать ДНК тест?
Помощь в развитии проекта

Помощь в развитии проекта
Ссылка на сбор
Гаплогруппа и субклад
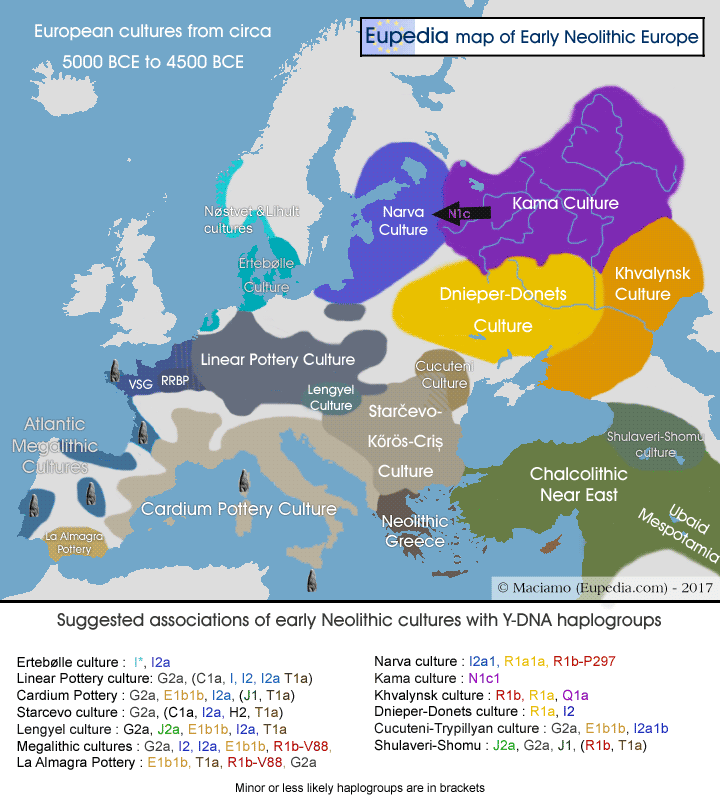
Исследовано 37 маркеров в лаборатории YSEQ
Гаплогруппа N1c. Наиболее глубокая прогнозная линия Y3037.
Полная линия:
Adam > A0-T > A1 > A1b > BT > CT > CF > F > GHIJK > HIJK > IJK > K > K2 > K-M2308 > K-M2335 > NO > N > Z4762 > L729 > F1360 > L666 > CTS4202 > Y3037
Указанная выше последовательность - это цепочка из субкладов, накопленных предками протестированного за более двухсоттысячелетний период, начиная с Y хромосомного Адама. Линии, находящиеся слева, являются предковыми к тем, что расположены правее.
Чем ближе к современности мы подбираемся, тем больше возможностей привязать линии к определённым этносам и субэтносам.
Первые 6 обозначений совпадают почти у всех не африканцев. У каждой ветки определен примерный возраст до последнего общего предка. Прародитель всех современных гаплогрупп кроме A и B жил около 68 тыс. лет назад. Эту линию называют макрогруппой (потому что она является предковой ко многим будущим гаплогруппам) CT.
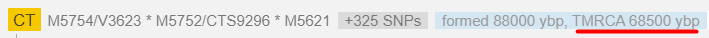
Общий предок всех кто относится к гаплогруппе N - 22 тыс. лет.
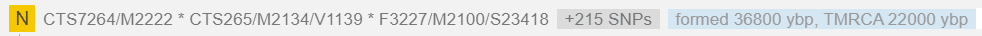
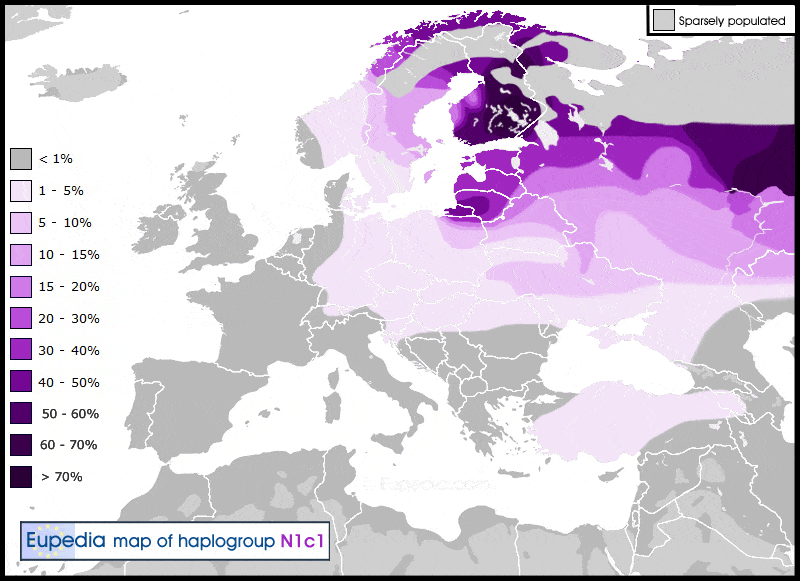
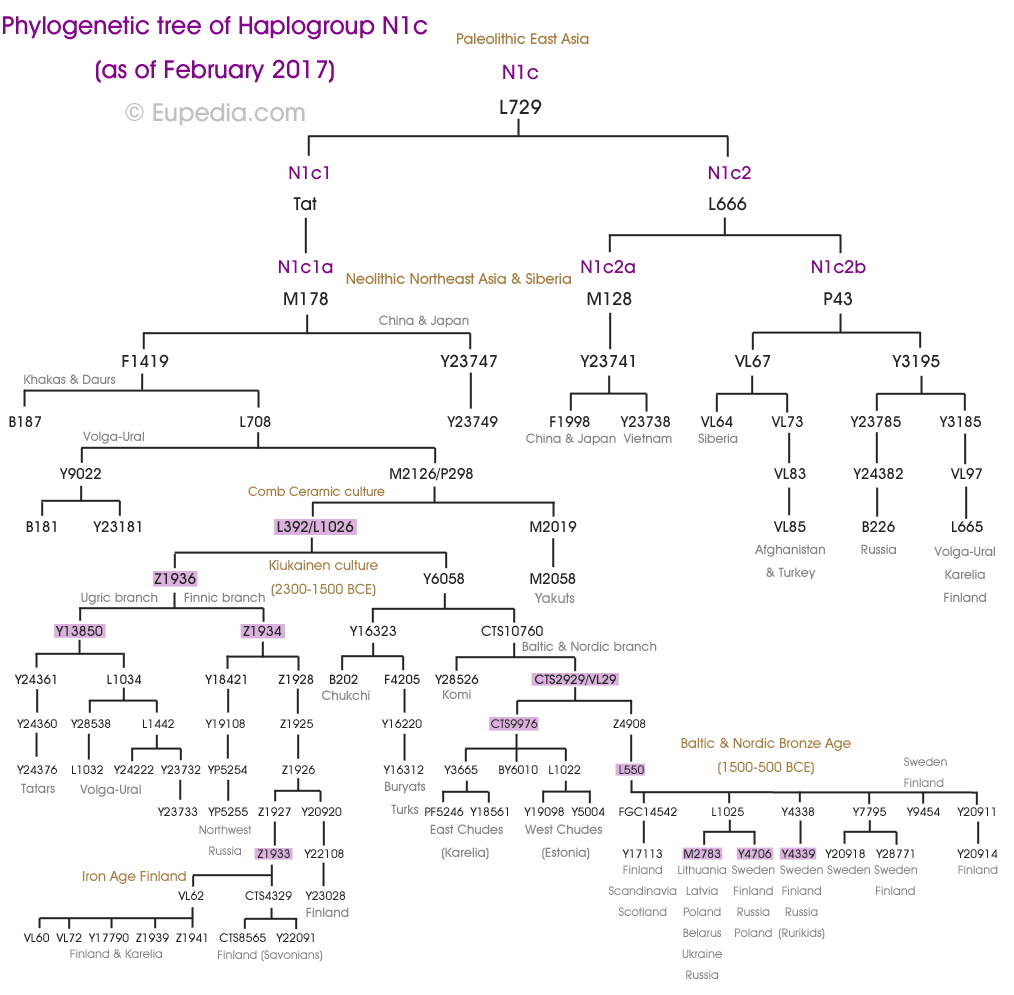
Гаплогруппа N является потомком восточноазиатской макрогаплогруппы NO. Считается, что она возникла в Индокитае или на юге Китая ~ 22 000 лет назад.
Гаплогруппы N1* и N1c были обнаружены с высокой частотой (26 из 70 образцов, или 37%) в останках эпохи неолита и бронзы (4500-700 гг. до н.э.) из долины реки Западный Ляо на северо-востоке Китая (Маньчжурия). Среди неолитических образцов гаплогруппа N1 составляла две трети образцов из культуры Хуншань (4700–2900 гг. до н. э.) и всех образцов из культуры Сяохэйан (3000–2200 гг. до н. э.), намекая на то, что N1 играли важную роль в распространении неолитического образа жизни вокруг Северо-Восточного Китая, а также, вероятно, в Монголию и Сибирь.
В 2016 г. было обнаружено 17 образцов представителей гапплогруппы N из культуры Сюэшань, датируемых 3600–2900 гг. до н.э., и среди них 41% принадлежал N1c1-Tat. Поэтому весьма вероятно, что субклад N1c1, обнаруженный сегодня в Европе, уходит своими корнями в китайский неолит. Он постепенно распространился по Сибири до северо-восточной Европы, возможно, достигнув Волго-Уральского региона около 5500–4500 гг. до н.э. с камской культурой (5300–3300 гг. до н.э.) и восточной Балтики с культурой гребенчатой керамики (4200–2000 гг. до н.э.). ), предполагаемая родовая культура протофинских и прибалтийских народов. Существует мало свидетельств земледелия или домашних животных в Сибири в эпоху неолита, но широко использовалась гончарная посуда. В этом отношении это было противоположное развитие Ближнего Востока, где сначала развилось сельское хозяйство, а затем только гончарное дело примерно с 5500 г. до н.э., возможно, благодаря контакту с выходцами из Восточной Азии через Сибирь или Среднюю Азию.
Образцы N1c найдены в культуре гребенчатой керамики на поздненеолитической стоянки Сертея II в Смоленской области России, недалеко от границы с Беларусью, которая датируется серединой 3-го тысячелетия до нашей эры.
Индоевропейская фатьяновско-балановская культура бронзового века (3200-2300 гг. до н.э.) постепенно захватила Балтийский регион и южную Финляндию с 2500 г. до н.э. (R1a ). Слияние двух групп, индоевропейской R1a и протоуральской N1c1, привело к возникновению гибридной культуры Киукайнен (2300–1500 гг. до н. э.). Современные балтийцы имеют примерно равную пропорцию гаплогрупп N1c1 и R1a, что является результатом слияния протоуральского и северо-восточного индоевропейского населения.
В 2018 году были протестированы шесть образцов возрастом 3500 лет с Кольского полуострова на северо-западе России и два образца идентифицированы как представители N1c-L392. Все они были аутосомно близки современным уральцам из Волго-Уральского региона и обладали типично уральскими линиями мтДНК (C4b, D4e4, T2d1b1, U4a1, U5a1d, Z1a1a).
Другое исследование показывает, что сибирская аутосомная ДНК и Y-гаплогруппа N1c отсутствовали в Эстонии бронзового века и не появлялись там до железного века, около 500 г. до н.э. Это показывает, что племена N1c сначала распространились на Беларусь, затем на Финляндию и на Кольский полуостров, прежде чем через несколько тысячелетий переселиться в Эстонию.
Филогения N1c1 показывает, что раскол между балто-финскими и уральскими (включая угорских) народами произошел около 4400 лет назад, после мутации L1026, почти точно в начале культуры Киукайнен. Уральская ветвь (Z1934) сформировалась первой около 4200 лет назад, за ней последовала угорская ветвь (Y13850) и, наконец, балто-финская ветвь (VL29) 3600 лет назад. Последние сразу разделились между чудями (CTS9976) на востоке и балто-финнами (L550) на западе. Фенноскандинавы (Y4706) и балты (M2783) раздвоились около 2600 лет назад.
Небольшой процент N1c1 встречается среди всего славянского, скандинавского населения, а также на большей части территории Германии (кроме северо-запада). Его происхождение в настоящее время неизвестно, но, скорее всего, он распространился с железным веком и раннесредневековыми (прото)славянскими племенами из России, Белоруссии и Украины в направлении Восточной Германии. Скандинавский N1c1 имеет три потенциальных источника:
- Прогрессивная ассимиляция северных саамов скандинавскими/германскими народами, начиная с железного века.
- Иммиграция из Германии и Польши в последние два тысячелетия.
- Обмен населением с Финляндией и странами Балтии, когда они попали под власть Скандинавии, особенно в течение восьми веков политического союза между Швецией и Финляндией.
Уральский N1c1
Гаплогруппа N1c1 тесно связана с уральскими народами, которые делятся на следующие семьи.
- Самодийские (нганасаны, энцы, ненцы и селькупы)
- финно-угорский
- финно-пермский
- Прибалтийско-финский (финский, карельский, эстонский и др.)
- Пермь (Коми, Удмуртия)
- Саамский (саамский)
- Волжский (марийский, мордовский)
- финно-пермский
- угорский
- венгерский язык
- Обско-угорские (ханты, манси)
Самоедская ветвь на севере Сибири раскололась раньше всех и соответствует субкладам N1c1* и N1c1a*.
Пермские и волжские носители имеют большое разнообразие субкладов N1c, включая N1c1a1 (L708), N1c1a1a (L1026), N1c1a1a1 (VL29), N1c1a1a2a (Z1935) и N1c2b (P43).
Балтийско-финская ветвь, по-видимому, возникла в результате миграции субклада N1c1a1a1 (VL29) из Волго-Уральского региона в Карелию, Финляндию и Эстонию. VL29 и его субклады также являются разновидностью N1c1, обнаруженной в балто-славянских популяциях, что подтверждает, что ветвь R1a индоевропейцев поглотила, а затем распространила линии N1c1 по Центральной и Восточной Европе.
Угорская ветвь, включающая венгерский язык, а также языки хантов и манси Западной Сибири, соответствует субкладу N1c1a1a2b (L1034).
Возникли ли урало-алтайские языки с гаплогруппой N в маньчжурском неолите?
Если гаплогруппа N1c, встречающаяся сегодня в основном среди носителей уральских языков, действительно возникла в маньчжурский неолит вместе с другими субкладами гаплогруппы N, это объясняет, что уральские языки родственны алтайским языкам, таким как тюркские и монгольские языки, а также более отдаленные корейский, японский языки, поскольку Y-гаплогруппа N действительно является объединяющим фактором между всеми этими популяциями. Некоторые лингвисты утверждают, что корейский, японский и айнский языки являются изолированными языками, поскольку они лишь отдаленно напоминают другие алтайские языки. Показательно, что в этих популяциях также самый низкий процент Y-гаплогруппы N — всего 3 % у корейцев и 2 % у японцев, в отличие от 10–25 % у монголов и бурят, и частоты от 30 до 90 % у большинства тюркоязычных сибиряков.
К известным представителям гаплогруппы N1c1 относятся:
Мономашичи (Рюриковичи) - Русские цари и основатель государственности на Руси.
Гедиминовичи - Великий Князь Литовский и его потомки Хованские, Голицыны, Трубецкие.
Гедройцы (Довспрунги) - Князь Литовский Гедрус и его потомки Гедройцы.
Линия предполагаемая у Крымукова чаще всего встречается в Восточной Азии (у китайцев и японцев). Также выявлена у татар. Среди других участников проекта совпадений нет, эта ветка крайне редко встречается на Кавказе.
Y-STR маркеры и Y-STR гаплотип
Подробнее о том, что такое Y-STR маркеры и Y-STR гаплотип можно почитать нажать на ссылку.
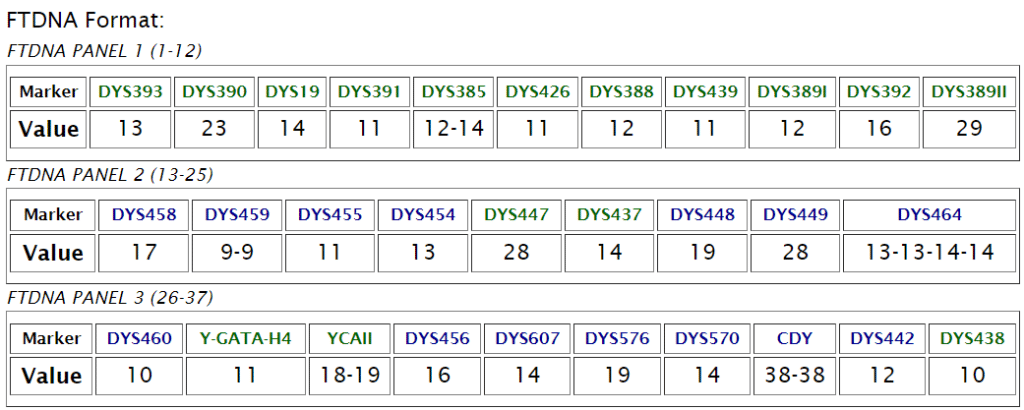
Nevgen
Nevgen - это онлайн-сервис, который позволяет предсказать Y-хромосомную гаплогруппу и субклад на основе STR-маркеров (коротких тандемных повторов). Он особенно полезен для тех, кто еще не сделал глубокий SNP-тест Y-хромосомы (BigY или аналоги), но уже имеет результаты по STR-маркерам (например, 12, 37, 67 или 111 маркеров).
Анализ STR-маркеров в сервисе Nevgen позволяет предсказать положение конкретной линии на дереве Y-хромосомных гаплогрупп и оценить целесообразность более глубокого SNP-тестирования
Подробнее по ссылке в шапке - Методика маркерного анализа
Калькулятор Nevgen не смог предсказать ни одну ветку в рамках гаплогруппы N. Очень редкое явление.
STR Match Finder
Видеоинструкция к STR Match Finder
STR Match Finder - это еще один полезный онлайн-инструмент для ДНК-генеалогии, который позволяет находить близких родственников по мужской линии на основе сравнения STR-маркеров Y-хромосомы. Он особенно актуален для тех, кто еще не сделал глубокий SNP-тест (BigY или аналоги), но хочет найти потенциальных родственников среди протестированных из той же популяции или гаплогруппы.
Поиск STR-совпадений в базе STR Match Finder помогает найти потенциальных родственников по мужской линии среди других протестированных из той же популяции. Мерой близости при этом служит генетическая дистанция, зависящая от числа мутационных различий
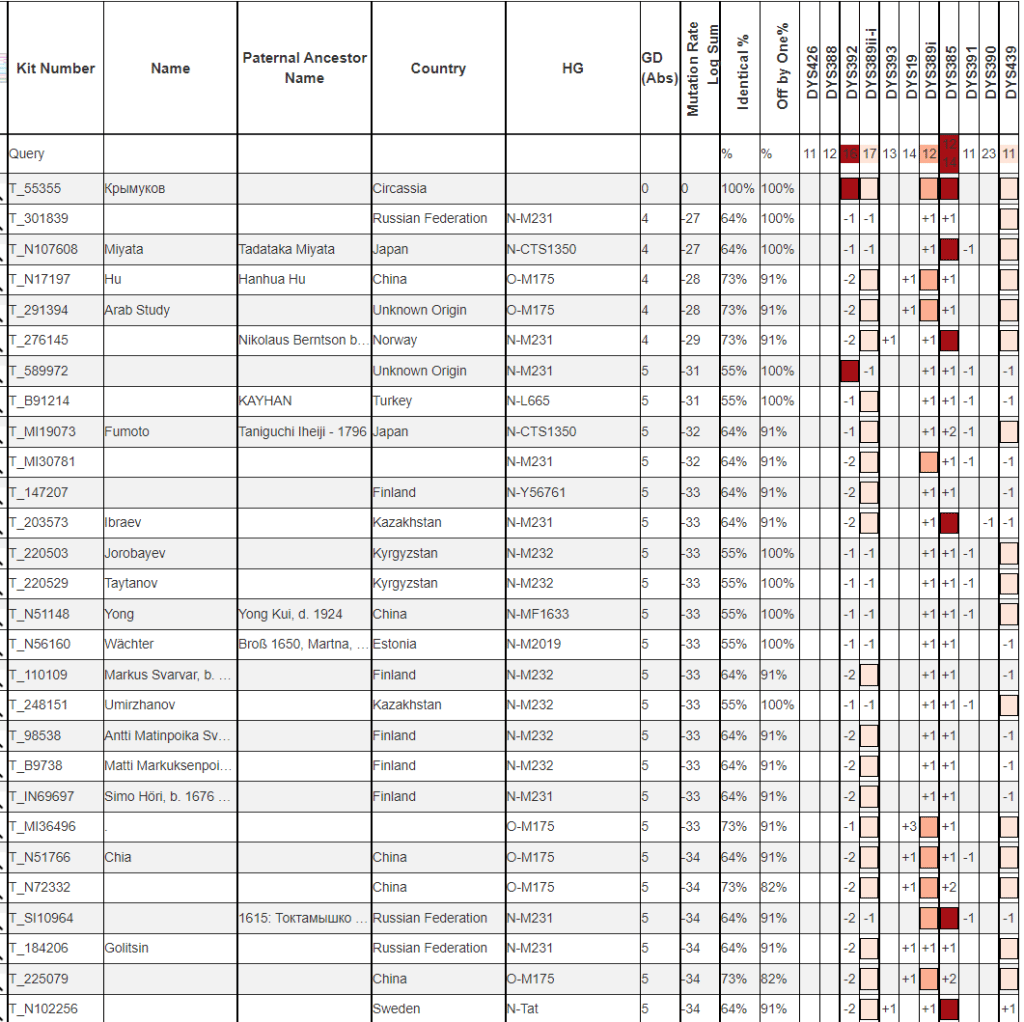
Очень далекие от всех результаты.
Y37
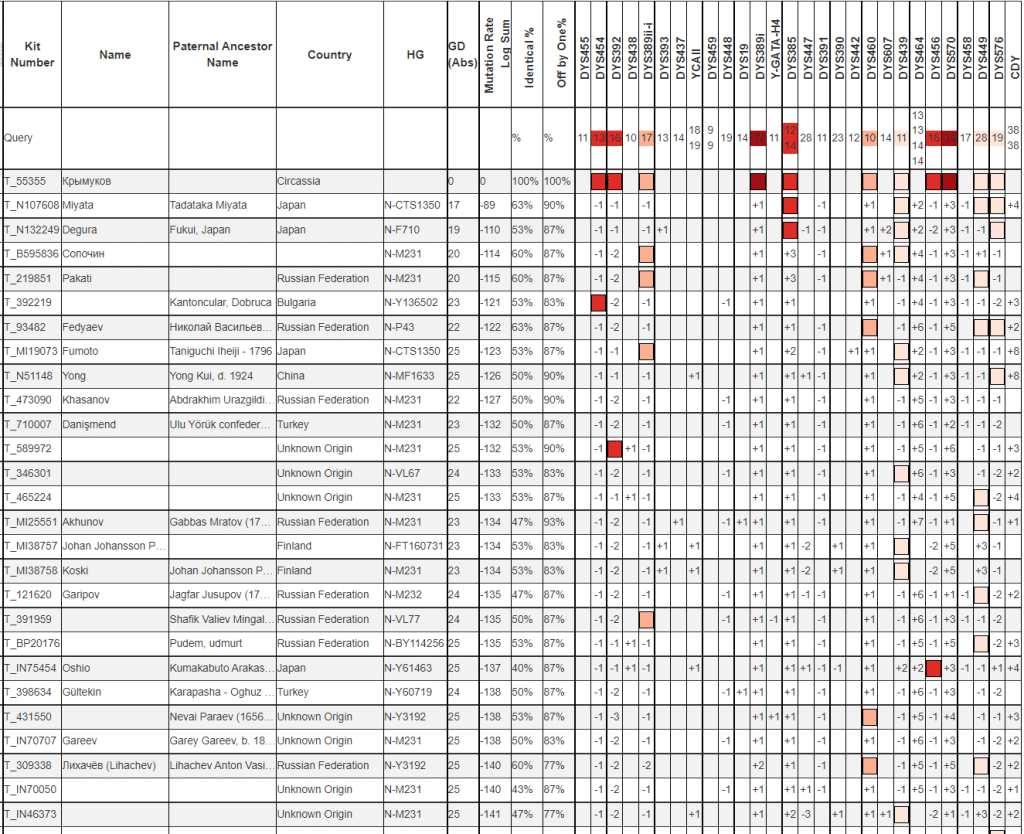
По 37 маркерам тоже очень большие дистанции. Если посмотреть на цифры в квадратиках, то заметно, что больше всего скачок цифр дают несколько маркеров в правой части таблицы. Посмотрим на совпадения без их учета.
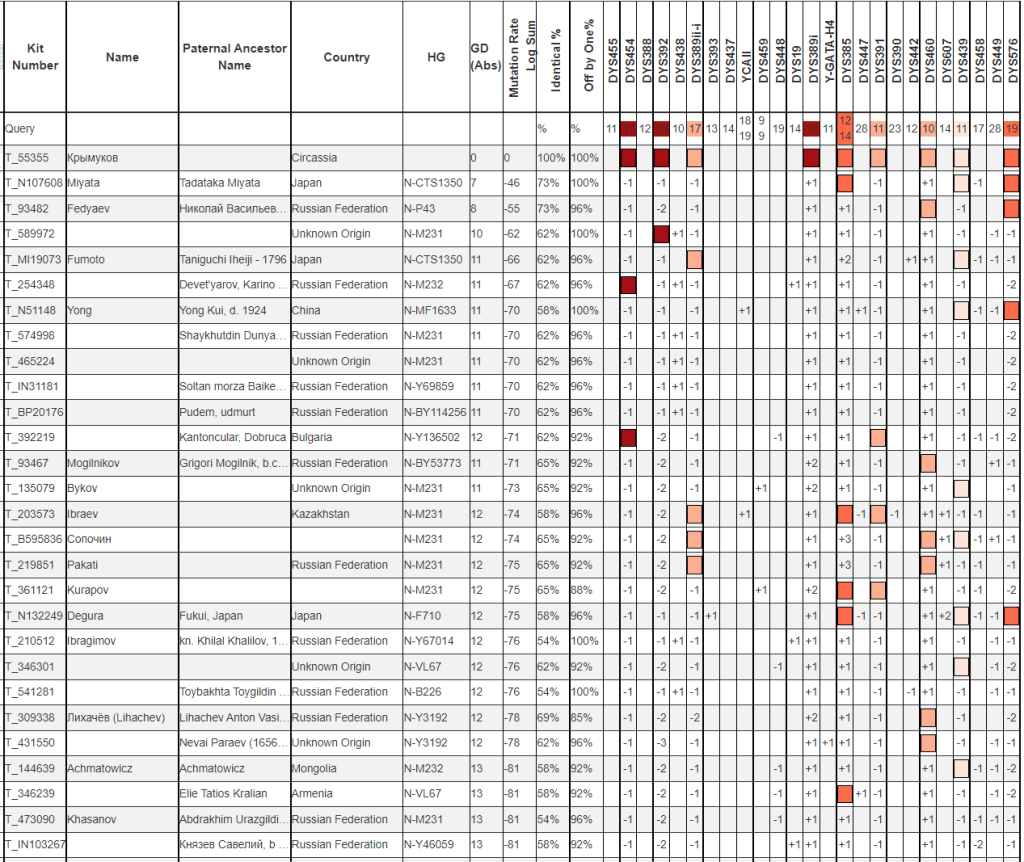
Даже с учетом коррекции картина не проясняется. Такие большие отличия явно свидетельствуют о том, что результат Крымукова будет явным образом относится к очень редким линиям гаплогруппы N. Единственный логический способ предсказания вероятной линии это подбор ближайших (даже если и не таких уж и близких). Большая часть из тех кто ближе к Крымукову относятся к линии N-Y3037.
Статистика проекта
По ссылке вы можете познакомится со статистикой. Сколько всего протестировано, информация по субэтносам, гаплогруппа итд.